
Строение цитоплазматической мембраны. Клеточная мембрана
Общие сведения об эукариотической клетке
Каждая эукариотическая клетка имеет обособленное ядро, в котором заключен отграниченный от матрикса ядерной мембраной генетический материал (это главное отличие от прокариотических клеток). Генетический материал сосредоточен преимущественно в виде хромосом, имеющих сложное строение и состоящих из нитей ДНК и белковых молекул. Деление клеток происходит посредством митоза (а для половых клеток - мейоза). Среди эукариотов есть как одноклеточные, так и многоклеточные организмы.
Существует несколько теорий происхождения эукариотических клеток, одна из них - эндосимбионтическая. В гетеротрофную анаэробную клетку проникла аэробная клетка типа бактериоподобной, которая послужила базой для появления митохондрий. В эти клетки начали проникать спирохетоподобные клетки, которые дали начало формированию центриолей. Наследственный материал отгородился от цитоплазмы, возникло ядро, появился митоз. В некоторые эукариотические клетки проникли клетки типа сине-зеленых водорослей, которые положили начало появлению хлоропластов. Так впоследствии возникло царство растений.
Размеры клеток тела человека варьируются от 2-7 мкм (у тромбоцитов) до гигантских размеров (до 140 мкм у яйцеклетки).
Форма клеток обусловлена выполняемой ими функцией: нервные клетки - звездчатые за счет большого количества отростков (аксона и дендритов), мышечные клетки - вытянутые, так как должны сокращаться, эритроциты могут менять свою форму при продвижении по мелким капиллярам.
Строение эукариотических клеток животных и растительных организмов во многом схоже. Каждая клетка снаружи ограничена клеточной оболочкой, или плазмалеммой. Она состоит из цито плазматической мембраны и слоя гликокаликса (толщиной 10-20 нм), который покрывает ее снаружи. Компоненты гликокаликса - комплексы полисахаридов с белками (гликопротеины) и жирами (гликолипиды).
Цитоплазматическая мембрана - это комплекс бислоя фосфолипидов с протеинами и полисахаридами.
В клетке выделяют ядро и цитоплазму . Клеточное ядро стоит из мембраны, ядерного сока, ядрышка и хроматина. Ядерная оболочка состоит из двух мембран, разделенных перинуклеарным пространством, и пронизана порами.
Основу ядерного сока (матрикса) составляют белки: нитчатые, мин фибриллярные (опорная функция), глобулярные, гетероядерные РНК и мРНК (результат процессинга).
Ядрышко - это структура, где происходит образование и созревание рибосомальных РНК (р-РНК).
Хроматин в виде глыбок рассеян в нуклеоплазме и является нитерфазной формой существования хромосом.
В цитоплазме выделяют основное вещество (матрикс, гиалоплазму), органеллы и включения.
Органеллы могут быть общего значения и специальные (в клетках, выполняющих специфические функции: микроворсинки всасывающего эпителия кишечника, миофибриллы мышечных клеток и т. д.).
Органеллы общего значения - эндоплазматическая сеть (гладкая и шероховатая), комплекс Гольджи, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра.
В растительных клетках есть еще и хлоропласты, в которых протекает фотосинтез.
Элементарная мембрана состоит из бислоя липидов в комплексе с белками (гликопротеины: белки + углеводы, липопротеины: жиры + белки). Среди липидов можно выделить фосфолипиды, холестерин, гликолипиды (углеводы + жиры), липопротеины. Каждая молекула жира имеет полярную гидрофильную головку и неполярный гидрофобный хвост. При этом молекулы ориентированы так, что головки обращены кнаружи и внутрь клетки, а неполярные хвосты - внутрь самой мембраны. Этим достигается избирательная проницаемость для веществ, поступающих в клетку.
Выделяют периферические белки (они расположены только по внутренней или наружной поверхности мембраны), интегральные (они прочно встроены в мембрану, погружены в нее, способны менять свое положение в зависимости от состояния клетки). Функции мембранных белков: рецепторная, структурная (поддерживают форму клетки), ферментативная, адгезивная, антигенная, транспортная.
Схема строения элементарной мембраны жидкостно-мозаичная: жиры составляют жидкокристаллический каркас, а белки мозаично встроены в него и могут менять свое положение.
Важнейшая функция: способствует компартментации - подразделению содержимого клетки на отдельные ячейки, отличающиеся деталями химического или ферментного состава. Этим достигается высокая упорядоченность внутреннего содержимого любой эукариотической клетки. Компартментация способствует пространственному разделению процессов, протекающих в клетке. Отдельный компартмент (ячейка) представлен какой-либо мембранной органеллой (например, лизосомой) или ее частью (кристами, отграниченными внутренней мембраной митохондрий).
Другие функции:
1) барьерная (отграничение внутреннего содержимого клетки);
2) структурная (придание определенной формы клеткам в соответствии с выполняемыми функциями);
3) защитная (за счет избирательной проницаемости, рецепции и антигенности мембраны);
4) регуляторная (регуляция избирательной проницаемости для различных веществ (пассивный транспорт без затраты энергии по законам диффузии или осмоса и активный транспорт с затратой энергии путем пиноцитоза, эндо- и экзоцитоза, работы натрий-калиевого насоса, фагоцитоза));
5) адгезивная функция (все клетки связаны между собой посредством специфических контактов (плотных и неплотных));
6) рецепторная (за счет работы периферических белков мембраны). Существуют неспецифические рецепторы, которые воспринимают несколько раздражителей (например, холодовые и тепловые терморецепторы), и специфические, которые воспринимают только один раздражитель (рецепторы световоспринимающей системы глаза);
7) электрогенная (изменение электрического потенциала поверхности клетки за счет перераспределения ионов калия и натрия ( мембранный потенциал нервных клеток составляет 90 мВ));
8) антигенная: связана с гликопротеинами и полисахаридами мембраны. На поверхности каждой клетки имеются белковые молекулы, которые специфичны только для данного вида клеток. С их помощью иммунная системы способна различать свои и чужие клетки.
Цитоплазма — обязательная часть клетки, заключенная между плазматической мембраной и ядром; подразделяется на гиалоплазму (основное вещество цитоплазмы), органоиды (постоянные компоненты цитоплазмы) и включения (временные компоненты цитоплазмы). Химический состав цитоплазмы: основу составляет вода (60-90% всей массы цитоплазмы), различные органические и неорганические соединения. Цитоплазма имеет щелочную реакцию. Характерная особенность цитоплазмы эукариотической клетки — постоянное движение ( циклоз ). Оно обнаруживается, прежде всего, по перемещению органоидов клетки, например хлоропластов. Если движение цитоплазмы прекращается, клетка погибает, так как, только находясь в постоянном движении, она может выполнять свои функции.
Гиалоплазма ( цитозоль ) представляет собой бесцветный, слизистый, густой и прозрачный коллоидный раствор. Именно в ней протекают все процессы обмена веществ, она обеспечивает взаимосвязь ядра и всех органоидов. В зависимости от преобладания в гиалоплазме жидкой части или крупных молекул, различают две формы гиалоплазмы: золь — более жидкая гиалоплазма и гель — более густая гиалоплазма. Между ними возможны взаимопереходы: гель превращается в золь и наоборот.
Функции цитоплазмы:
- объединение всех компонентов клетки в единую систему,
- среда для прохождения многих биохимических и физиологических процессов,
- среда для существования и функционирования органоидов.
Клеточные оболочки
Клеточные оболочки ограничивают эукариотические клетки. В каждой клеточной оболочке можно выделить как минимум два слоя. Внутренний слой прилегает к цитоплазме и представлен плазматической мембраной (синонимы — плазмалемма, клеточная мембрана, цитоплазматическая мембрана), над которой формируется наружный слой. В животной клетке он тонкий и называется гликокаликсом (образован гликопротеинами, гликолипидами, липопротеинами), в растительной клетке — толстый, называется клеточной стенкой (образован целлюлозой).
Все биологические мембраны имеют общие структурные особенности и свойства. В настоящее время общепринята жидкостно-мозаичная модель строения мембраны . Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Фосфолипиды — триглицериды, у которых один остаток жирной кислоты замещен на остаток фосфорной кислоты; участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой, участки, в которых находятся остатки жирных кислот — гидрофобными хвостами. В мембране фосфолипиды располагаются строго упорядоченно: гидрофобные хвосты молекул обращены друг к другу, а гидрофильные головки — наружу, к воде.
Помимо липидов в состав мембраны входят белки (в среднем ≈ 60%). Они определяют большинство специфических функций мембраны (транспорт определенных молекул, катализ реакций, получение и преобразование сигналов из окружающей среды и др.). Различают: 1) периферические белки (расположены на наружной или внутренней поверхности липидного бислоя), 2) полуинтегральные белки (погружены в липидный бислой на различную глубину), 3) интегральные, или трансмембранные, белки (пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки). Интегральные белки в ряде случаев называют каналообразующими, или канальными, так как их можно рассматривать как гидрофильные каналы, по которым в клетку проходят полярные молекулы (липидный компонент мембраны их бы не пропустил).

А — гидрофильная головка фосфолипида; В — гидрофобные хвостики фосфолипида; 1 — гидрофобные участки белков Е и F; 2 — гидрофильные участки белка F; 3 — разветвленная олигосахаридная цепь, присоединенная к липиду в молекуле гликолипида (гликолипиды встречаются реже, чем гликопротеины); 4 — разветвленная олигосахаридная цепь, присоединенная к белку в молекуле гликопротеина; 5 — гидрофильный канал (функционирует как пора, через которую могут проходить ионы и некоторые полярные молекулы).
В состав мембраны могут входить углеводы (до 10%). Углеводный компонент мембран представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Углеводы обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину несколько десятков нанометров. В нем располагаются многие рецепторы клетки, с его помощью происходит адгезия клеток.
Молекулы белков, углеводов и липидов подвижны, способны перемещаться в плоскости мембраны. Толщина плазматической мембраны — примерно 7,5 нм.
Функции мембран
Мембраны выполняют такие функции:
- отделение клеточного содержимого от внешней среды,
- регуляция обмена веществ между клеткой и средой,
- деление клетки на компартаменты («отсеки»),
- место локализации «ферментативных конвейеров»,
- обеспечение связи между клетками в тканях многоклеточных организмов (адгезия),
- распознавание сигналов.
Важнейшее свойство мембран — избирательная проницаемость, т.е. мембраны хорошо проницаемы для одних веществ или молекул и плохо проницаемы (или совсем непроницаемы) для других. Это свойство лежит в основе регуляторной функции мембран, обеспечивающей обмен веществ между клеткой и внешней средой. Процесс прохождения веществ через клеточную мембрану называют транспортом веществ . Различают: 1) пассивный транспорт — процесс прохождения веществ, идущий без затрат энергии; 2) активный транспорт — процесс прохождения веществ, идущий с затратами энергии.
При пассивном транспорте вещества перемещаются из области с более высокой концентрацией в область с более низкой, т.е. по градиенту концентрации. В любом растворе имеются молекулы растворителя и растворенного вещества. Процесс перемещения молекул растворенного вещества называют диффузией, перемещения молекул растворителя — осмосом. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Можно выделить следующие виды пассивного транспорта: 1) простая диффузия — транспорт веществ непосредственно через липидный бислой (кислород, углекислый газ); 2) диффузия через мембранные каналы — транспорт через каналообразующие белки (Na + , K + , Ca 2+ , Cl -); 3) облегченная диффузия — транспорт веществ с помощью специальных транспортных белков, каждый из которых отвечает за перемещение определенных молекул или групп родственных молекул (глюкоза, аминокислоты, нуклеотиды); 4) осмос — транспорт молекул воды (во всех биологических системах растворителем является именно вода).
Необходимость активного транспорта возникает тогда, когда нужно обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется особыми белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ. К активному транспорту относят: 1) Na + /К + -насос (натрий-калиевый насос), 2) эндоцитоз, 3) экзоцитоз.

Работа Na + /К + -насоса . Для нормального функционирования клетка должна поддерживать определенное соотношение ионов К + и Na + в цитоплазме и во внешней среде. Концентрация К + внутри клетки должна быть значительно выше, чем за ее пределами, а Na + — наоборот. Следует отметить, что Na + и К + могут свободно диффундировать через мембранные поры. Na + /К + -насос противодействует выравниванию концентраций этих ионов и активно перекачивает Na + из клетки, а K + в клетку. Na + /К + -насос представляет собой трансмембранный белок, способный к конформационным изменениям, вследствие чего он может присоединять как K + , так и Na + . Цикл работы Na + /К + -насоса можно разделить на следующие фазы: 1) присоединение Na + с внутренней стороны мембраны, 2) фосфорилирование белка-насоса, 3) высвобождение Na + во внеклеточном пространстве, 4) присоединение K + с внешней стороны мембраны, 5) дефосфорилирование белка-насоса, 6) высвобождение K + во внутриклеточном пространстве. На работу натрий-калиевого насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки. За один цикл работы насос выкачивает из клетки 3Na + и закачивает 2К + .
Эндоцитоз — процесс поглощения клеткой крупных частиц и макромолекул. Различают два типа эндоцитоза: 1) фагоцитоз — захват и поглощение крупных частиц (клеток, частей клеток, макромолекул) и 2) пиноцитоз — захват и поглощение жидкого материала (раствор, коллоидный раствор, суспензия). Явление фагоцитоза открыто И.И. Мечниковым в 1882 г. При эндоцитозе плазматическая мембрана образует впячивание, края ее сливаются, и происходит отшнуровывание в цитоплазму структур, отграниченных от цитоплазмы одиночной мембраной. К фагоцитозу способны многие простейшие, некоторые лейкоциты. Пиноцитоз наблюдается в эпителиальных клетках кишечника, в эндотелии кровеносных капилляров.
Экзоцитоз — процесс, обратный эндоцитозу: выведение различных веществ из клетки. При экзоцитозе мембрана пузырька сливается с наружной цитоплазматической мембраной, содержимое везикулы выводится за пределы клетки, а ее мембрана включается в состав наружной цитоплазматической мембраны . Таким способом из клеток желез внутренней секреции выводятся гормоны, у простейших — непереваренные остатки пищи.
Перейти к лекции №5 «Клеточная теория. Типы клеточной организации»
Перейти к лекции №7 «Эукариотическая клетка: строение и функции органоидов»
Развивался таким образом, что функция каждой его системы стала результатом функции суммы клеток, из которых состоят органы и ткани данной системы. Каждая клетка организма располагает набором структур и механизмов, позволяющих ей осуществлять собственный метаболизм и выполнять присущую ей функцию.
В состав клетки входят цитоплазматическая или поверхностная мембрана; цитоплазма, имеющая ряд органелл, включений, элементов цитоскелета; ядро, содержащее ядерный геном. Органеллы клетки и ядро отграничены в цитоплазме внутренними мембранами. Каждая структура клетки выполняет в ней свою функцию, а все они вместе взятые обеспечивают жизнеспособность клетки и выполнение ею специфических функций.
Ключевая роль в осуществлении клеточных функций и их регуляции принадлежит цитоплазматической мембране клетки.
Общие принципы строения цитоплазматической мембраны
Для всех клеточных мембран характерен один принцип строения (рис. 1), в основе которого лежат физико-химические свойства сложных липидов и белков, входящих в их состав. Мембраны клетки располагаются в водной среде и для понимания физико-химических явлений, влияющих на их структурную организацию , полезным является описание взаимодействия липидных и белковых молекул с молекулами воды и друг с другом. Ряд свойств клеточных мембран также вытекает из рассмотрения этого взаимодействия.
Известно, что плазматическая мембрана клетки представлена двойным слоем сложных липидов, покрывающим поверхность клетки на всем ее протяжении. Для создания липидного бислоя в ее структуру могли быть отобраны природой и включены только те молекулы липидов, которые обладают амфифильными (амфипатическими) свойствами. Этим условиям отвечают молекулы фосфолипидов и холестерола. Их свойства таковы, что одна часть молекулы (глицерольная для фосфолипидов и циклопентановая для холестерола) обладает полярными (гидрофильными) свойствами, а другая (жирнокислотные радикалы) — неполярными (гидрофобными) свойствами.
Рис. 1. Строение цитоплазматической мембраны клетки.
Если определенное количество молекул фосфолипидов и холестерола поместить в водную среду, то они спонтанно начнут собираться в упорядоченные структуры и формировать замкнутые пузырьки ( липосомы ), в которых оказывается заключенной часть водной среды, а поверхность становится покрытой непрерывным двойным слоем ( бислоем ) фосфолипидных молекул и холестерола. При рассмотрении характера пространственного расположения молекул фосфолипидов и холестерола в этом бислое видно, что молекулы данных веществ располагаются своими гидрофильными частями в сторону наружного и внутреннего водных пространств, а гидрофобными — в противоположных направлениях — внутрь бислоя.
Что заставляет молекулы этих липидов самопроизвольно формировать в водной среде бислойные структуры, подобные структуре бислоя клеточной мембраны? Пространственное расположение амфифильных молекул липидов в водной среде диктуется одним из требований термодинамики. Наиболее вероятной пространственной структурой, которую сформируют в водной среде молекулы липидов, будет структура, обладающая минимумом свободной энергии .
Такой минимум свободной энергии в пространственной структуре липидов в воде будет достигнут в случае, когда и гидрофильные, и гидрофобные свойства молекул будут реализованы в виде соответствующих межмолекулярных связей.
При рассмотрении поведения сложных амфифильных молекул липидов в воде можно объяснить и некоторые свойства клеточных мембран . Известно, что если механически повредить плазматическую мембрану (например, проколоть ее электродом или через прокол удалить ядро и поместить в клетку другое ядро), то через мгновение за счет сил межмолекулярного взаимодействия липидов и воды мембрана самопроизвольно восстановит целостность . Под действием таких же сил можно наблюдать слияние бислоев двух мембран при их соприкосновении (например, везикул и пресинаптической мембраны в синапсах). Способность мембран сливаться при их непосредственном контакте является частью механизмов обновления структуры мембран, транспорта компонентов мембран из одного субклеточного пространства в другое, а также частью механизмов эндо- и экзоцитоза.
Энергия межмолекулярных связей в липидном бислое очень низкая, поэтому создаются условия для быстрого перемещения в мембране молекул липидов и белков и для изменения структуры мембраны при воздействии на нее механических сил , давлений, температуры и других факторов. Наличие в мембране двойного липидного слоя образует замкнутое пространство, изолирует цитоплазму от окружающей водной среды и создаст препятствие для свободного прохождения воды и растворимых в ней веществ через клеточную мембрану. Толщина липидного бислоя составляет около 5 нм.
В состав клеточных мембран также входят белки. Их молекулы по объему и массе в 40-50 раз больше, чем молекулы мембранных липидов. За счет белков толщина мембраны достигает 7-10 нм. Несмотря на то что суммарные массы белков и липидов в большинстве мембран почти равны, количество молекул белков в мембране в десятки раз меньше, чем молекул липидов.
Что же произойдет, если белковая молекула окажется помещенной в фосфолипидный бислой липосом, наружные и внутренние поверхности которых полярны, а внутрилипидный неполярен? Под влиянием сил межмолекулярных взаимодействий липидов, белка и воды произойдет формирование такой пространственной структуры, в которой неполярные участки пептидной цепи будут стремиться расположиться в глубине липидного бислоя, в то время как полярные — занять положение на одной из поверхностей бислоя и могут к тому же оказаться погруженными во внешнюю или внутреннюю водную среду липосомы. Очень сходный характер расположения белковых молекул имеет место и в липидном бислое клеточных мембран (рис. 1).
Обычно белковые молекулы локализуются в мембране разрозненно одна от другой. Возникающие в неполярной части бислоя липидов очень слабые силы гидрофобных взаимодействий между углеводородными радикалами липидных молекул и неполярными участками белковой молекулы (липид-липидные, липид-белковые взаимодействия) не препятствуют протеканию процессов тепловой диффузии этих молекул в структуре бислоя.
Когда с помощью тонких методов исследования была изучена структура клеточных мембран, то оказалось, что она очень сходна с той, которая самопроизвольно формируется фосфолипидами, холестеролом и белками в водной среде. В 1972 г. Синджером и Никольсом была предложена жидкостно-мозаичная модель строения клеточной мембраны и сформулированы ее основные принципы.
Согласно этой модели, структурную основу всех клеточных мембран составляет жидкоподобный непрерывный двойной слой амфипатических молекул фосфолипидов, холестсрола, гликолипидов, самопроизвольно формирующих его в водной среде. В липидном бислое асимметрично расположены белковые молекулы, выполняющие специфические рецепторные, ферментативные и транспортные функции. Белковые и липидные молекулы обладают подвижностью и могут совершать вращательные движения , диффундировать в плоскости бислоя. Белковые молекулы способны изменять их пространственную структуру (конформацию), смещаться и изменять свое положение в липидном бислое мембраны, погружаясь на различную глубину или всплывая на его поверхность. Структура липидного бислоя мембраны неоднородна. В нем имеются участки (домены), получившие название «рафты», которые обогащены сфинголипидами и холестеролом. «Рафты» отличаются фазовым состоянием от состояния остальной части мембраны, в которой они располагаются. Особенности строения мембран зависят от выполняемой ими функции и функционального состояния.
Исследование состава клеточных мембран подтвердили, что основными их компонентами являются липиды, составляющие около 50% массы плазматической мембраны. Около 40-48% массы мембраны приходится на белки и 2-10% — на углеводы. Остатки углеводов либо входят в состав белков, образуя гликопротеины, либо липидов, образуя гликолипиды. Фосфолипиды являются главными структурными липидами плазматических мембран и составляют 30-50% их массы.
Углеводные остатки молекул гликолипидов обычно располагаются на внешней поверхности мембраны и погружены в водную среду. Они играют важную роль в межклеточных, клеточно-матриксных взаимодействиях и распознавании антигенов клетками иммунной системы. Молекулы холестерола, встроенные в фосфолипидный бислой, способствуют сохранению упорядоченного расположения жирнокислотных цепей фосфолипидов и их жидкокристаллического состояния. В связи с наличием высокой конформационной подвижности ацильных радикалов жирных кислот фосфолипидов они формируют достаточно рыхлую упаковку липидного бислоя и в нем могут образовываться структурные дефекты.
Белковые молекулы способны пронизывать всю мембрану так, что их концевые участки выступают за се поперечные пределы. Такие белки называют трансмембранными , или интегральными . В составе мембран имеются также белки, только частично погруженные в мембрану или располагающиеся на ее поверхности.
Многие специфические функции мембран определяются белковыми молекулами, для которых липидная матрица является непосредственным микроокружением и от ее свойств зависит осуществление функций белковыми молекулами. Среди важнейших функций мембранных белков можно выделить: рецепторную — связывание с такими сигнальными молекулами, как нейромедиаторы, гормоны, ингерлейкины, факторы роста, и передача сигнала на пострецепторные структуры клетки; ферментативную — катализ внутриклеточных реакций; структурную — участие в формировании структуры самой мембраны; транспортную — перенос веществ через мембраны; каналообразующую — формирование ионных и водных каналов. Белки совместно с углеводами участвуют в осуществлении адгезии-слипания, склеивания клеток при иммунных реакциях, объединении клеток в слои и ткани, обеспечивают взаимодействие клеток с внеклеточным матриксом.
Функциональная активность мембранных белков (рецепторов, ферментов, переносчиков) определяется их способностью легко изменять свою пространственную структуру (конформацию) при взаимодействии с сигнальными молекулами, действии физических факторов или изменении свойств среды микроокружения. Энергия, требующаяся для осуществления этих конформационных изменений структуры белков, зависит как от внутримолекулярных сил взаимодействия отдельных участков пептидной цепи, так и от степени текучести (микровязкости) мембранных липидов, непосредственно окружающих белок.
Углеводы в виде гликолипидов и гликопротеинов составляют лишь 2-10% от массы мембраны; количество их в разных клетках изменчиво. Благодаря им осуществляются некоторые виды межклеточных взаимодействий, они принимают участие в узнавании клеткой чужеродных антигенов и совместно с белками создают своеобразную антигенную структуру поверхностной мембраны собственной клетки. По таким антигенам клетки узнают друг друга, объединяются в ткань и на короткое время слипаются для передачи сигнальных молекул друг другу.
Благодаря низкой энергии взаимодействия входящих в мембрану веществ и относительной упорядоченности их расположения клеточная мембрана приобретает ряд свойств и функций, не сводимых к простой сумме свойств образующих ее веществ. Незначительные по силе воздействия на мембрану, сравнимые с энергией межмолекулярных связей белков и липидов, могут вести к изменению конформации белковых молекул, проницаемости ионных каналов, изменению свойств мембранных рецепторов, других многочисленных функций мембраны и самой клетки. Высокая чувствительность структурных компонентов плазматической мембраны имеет решающее значение в восприятии клеткой информационных сигналов и их преобразовании в ответные реакции клетки.
Функции цитоплазматической мембраны клетки
Цитоплазматическая мембрана выполняет многие функции, обеспечивающие жизненные потребности клетки и, в частности, ряд функций необходимых для восприятия и передачи клеткой информационных сигналов.
Среди важнейших функций плазматической мембраны можно выделить:
- отграничение клетки от окружающей се среды с сохранением формы, объема и существенных различий между клеточным содержимым и внеклеточным пространством;
- перенос веществ внутрь клетки и из нее на основе свойства избирательной проницаемости, активного и других видов транспорта;
- поддержание трансмембранной разности электрических потенциалов (поляризации мембраны) в покое, ее изменение при различных воздействиях на клетку, генерация и проведение возбуждения;
- участие в обнаружении (рецепции) сигналов физической природы , сигнальных молекул за счет формирования сенсорных или молекулярных рецепторов и передаче сигналов внутрь клетки;
- образование межклеточных контактов (плотный, щелевой и десмосомальный контакт) в составе образуемых тканей или при адгезии клеток различных тканей;
- создание гидрофобного микроокружения для проявления активности ферментов, связанных с мембраной;
- обеспечение иммунной специфичности клетки за счет наличия в структуре мембраны антигенов белковой или гликопротеиновой природы. Иммунная специфичность имеет значение при объединении клеток в ткань и взаимодействии с клетками, осуществляющими иммунный надзор в организме.
Приведенный перечень функций клеточных мембран свидетельствует о том, что они принимают участие в осуществлении не только клеточных функций, но и базисных процессов жизнедеятельности органов, тканей и целостного организма. Без знания ряда явлений и процессов, обеспечиваемых мембранными структурами, невозможно понимание и осознанное выполнение некоторых диагностических процедур и лечебных мероприятий. Например, для правильного применения многих лекарственных веществ необходимо знание того, в какой мере каждое из них проникает через клеточные мембраны из крови в тканевую жидкость и в клетки.
Наружная цитоплазматическая мембрана представляет собой тончайшую пленку. Ее толщина - порядка 7-10 нм. Просматривается пленка только в электронный микроскоп.
Структура
Какой состав имеет цитоплазматическая мембрана? Строение пленки достаточно разнообразно. В соответствии с химической организацией, она представляет собой комплекс белков и липидов. Цитоплазматическая мембрана клетки включает в себя бислой. Он выступает в качестве основы. Кроме этого, цитоплазматическая мембрана содержит холестерол и гликолипиды. Этим веществам свойственна амфипатричность. Другими словами, в них присутствуют гидрофобные ("боящиеся влаги") и гидрофильные ("любящие воду") концы. Последние (фосфатная группа) направлены наружу от мембраны, вторые (остатки от жирных кислот) ориентированы друг к другу. За счет этого и формируется липидный биполярный слой. Липидные молекулы обладают подвижностью. Они способны перемещаться в собственном монослое либо (что редко) из одного в другой.
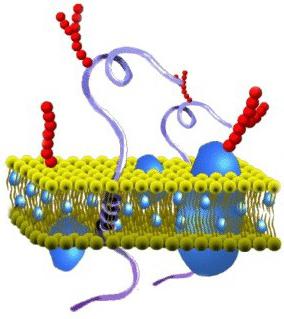
Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды - основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Цитоплазматическая мембрана: функции
Основными задачами являются:
- Барьерная . Защитная пленка обеспечивает активный, пассивный, избирательный, регулируемый обмен соединений с внешней средой. За счет избирательной проницаемости осуществляется отделение клетки и ее компартментов и снабжение их нужными веществами.
- Транспортная . Сквозь пленку осуществляется переход соединений от клетки к клетке. Благодаря этому доставляются питательные соединения, удаляются конечные продукты обмена, происходит секреция разных веществ. Кроме этого, формируются ионные градиенты, на оптимальном уровне поддерживаются ионная концентрация и рН. Они необходимы для активной деятельности ферментов клетки.
Вспомогательные задачи
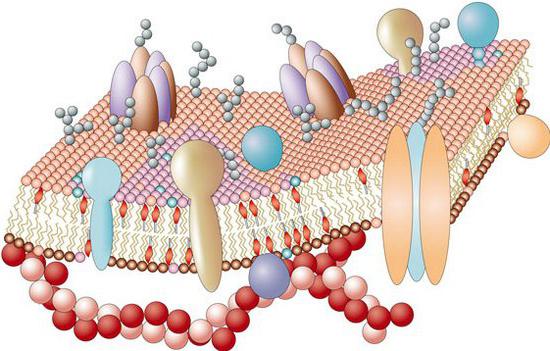
Особые свойства
К специфическим функциям мембраны относят:

Дополнительные сведения
Если какие-то частицы по тем или другим причинам не способны пройти сквозь фосфолипидный бислой (к примеру, вследствие гидрофильных свойств, поскольку внутри цитоплазматическая мембрана гидрофобна и такие соединения не пропускает, либо из-за больших размеров самих частиц), но они необходимы, то пройти они могут с помощью специальных белков-переносчиков (транспортеров) и белков-каналов. Либо проникновение их осуществляется посредством эндоцитоза.
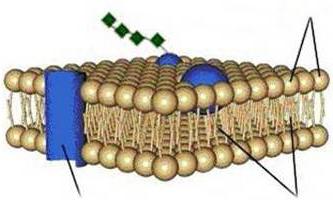
В процессе пассивного транспорта пересечение веществами липидного слоя происходит путем диффузии. При этом энергия не затрачивается. В качестве одного из вариантов такого механизма может выступать облегченная диффузия. В ходе нее облегчает прохождение вещества какая-нибудь специфическая молекула. У нее может присутствовать канал, способный пропускать только однотипные частицы. При активном транспорте затрачивается энергия. Это связано с тем, что данный процесс осуществляется против концентрационного градиента. Цитоплазматическая мембрана содержит особые белки-насосы, АТФазу в том числе, которая способствует активному вхождению калиевых и выведению натриевых ионов.
Модели
Их существует несколько:
- "Бутербродная модель" . Идею о трехслойном строении всех мембран высказали ученые Даусон и Даниэли в 1935 году. По их мнению, структура пленки была следующей: белки-липиды-белки. Такое представление существовало достаточно долго.
- "Жидкостно-мозаичная структура". Эта модель была описана Николсоном и Сингером в 1972 году. В соответствии с ней белковые молекулы не формируют сплошной слой, а погружаются в биполярный липидный в виде мозаики на различную глубину. Эта модель считается наиболее универсальной.
- "Белково-кристаллическая структура". В соответствии с этой моделью мембраны формируются за счет переплетения белковых и липидных молекул, которые объединены на базе гидрофильно-гидрофобных связей.
Строение клетки
Современное определение клетки следующее: клетка - это открытая, ограниченная активной мембраной, структурированная система биополимеров (белков и нуклеиновых кислот) и их макромолекулярных комплексов, участвующих в единой совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Есть и другое определение клетки. Клетка - это возникшая в результате эволюции, открытая биологическая система , ограниченная полупроницаемой мембраной, состоящая из ядра и цитоплазмы, способная к саморегуляции и самовоспроизведению.
Как мы видим из определений, устройство клетки достаточно сложно. Кроме того, говоря о клетках, мы можем иметь в виду клетки разных организмов, тканей органов. Таким образом, каждая разновидность клетки имеет свои неповторимые особенности. Давайте постараемся выбрать из этого разнообразия те черты и особенности, которые объединяют клетки разных типов. Идеальная клетка состоит из трех частей: клеточной мембраны, ядра, цитоплазмы с органоидами и органеллами.
Цитоплазматическая мембрана (ЦПМ)
Строение мембраны во многом остается загадочным. Существовало несколько теорий относительно строения ПМ. Еще в 30-х годах ХХ века была выдвинута гипотеза, названная по имени её авторов моделью Давсона – Данеэли (модель сендвича или бутербродная модель). По этой модели в основе мембраны лежит двойной гидрофобный слой жиров. Этот слой окружен двумя слоями белков.
Однако к началу 70-х годов XX века накопились данные, противоречащие этой гипотезе. В результате была выдвинута модель, получившая название модели Сингера – Николсона. Это модель динамической мембраны. В основе этой модели все тот же двойной слой жиров, но белки, согласно этой модели подвижные острова в море жиров.
Клеточная (или плазматическая) мембрана представляет собой тонкую, гибкую и эластичную структуру толщиной всего 7,5-10 нм. Она состоит в основном из белков и липидов. Примерное соотношение ее компонентов таково: белки - 55%, фосфолипиды - 25%, холестерол - 13%, другие липиды - 4%, углеводы - 3%.
Липидный слой клеточной мембраны препятавует проникновению воды. Основу мембраны составляет липидный бислой - тонкая липидная пленка, состоящая из двух монослоев и полностью покрывающая клетку. По всей мембране располагаются белки в виде крупных глобул.
Липидный бислой состоит главным образом из молекул фосфолипидов . Один конец такой молекулы является гидрофильным, т.е. растворимым в воде (на нем расположена фосфатная группа), другой - гидрофобным, т.е. растворимым только в жирах (на нем находится жирная кислота).
Благодаря тому, что гидрофобная часть молекулы фосфолипида отталкивает воду, но притягивается к подобным частям таких же молекул, фосфолипиды имеют природное свойство прикрепляться друг к другу в толще мембраны. Гидрофильная часть с фосфатной группой образует две мембранные поверхности: наружную, которая контактирует с внеклеточной жидкостью, и внутреннюю, которая контактирует с внутриклеточной жидкостью.
Середина липидного слоя непроницаема для ионов и водных растворов глюкозы и мочевины. Жирорастворимые вещества, включая кислород, углекислый газ, алкоголь, напротив, легко проникают через эту область мембраны.
Молекулы холестерола , входящего в состав мембраны, по природе также относятся к липидам, поскольку их стероидная группировка обладает высокой растворимостью в жирах. Эти молекулы как бы растворены в липидном бислое. Их главное назначение - регуляция проницаемости (или непроницаемости) мембран для водорастворимых компонентов жидких сред организма. Кроме того, холестерол - основной регулятор вязкости мембраны.
Белки клеточных мембран. На рисунке в липидном бислое видны глобулярные частицы - это мембранные белки, большинство которых являются гликопротеинами. Различают два типа мембранных белков: (1) интегральные, которые пронизывают мембрану насквозь; (2) периферические, которые выступают только над одной ее поверхностью, не достигая другой.
Многие интегральные белки формируют каналы (или поры), через которые во внутри- и внеклеточную жидкость могут диффундировать вода и водорастворимые вещества, особенно ионы. Благодаря избирательности действия каналов одни вещества диффундируют лучше других.
Другие интегральные белки функционируют как белки-переносчики, осуществляя транспорт веществ, для которых липидный бислой непроницаем. Иногда белки-переносчики действуют в направлении, противоположном диффузии, такой транспорт называют активным. Некоторые интегральные белки являются ферментами.
Интегральные белки мембраны могут служить также рецепторами для водорастворимых веществ, включая пептидные гормоны, поскольку мембрана для них непроницаема. Таким образом, интегральные белки, встроенные в клеточную мембрану, вовлекают ее в процесс передачи информации о внешней среде внутрь клетки.
Плазматическая мембрана, или плазмалемма, ограничивает клетку снаружи, выполняя роль механического барьера. Через нее происходит транспорт веществ внутрь клетки и наружу. Мембрана обладает свойством избирательной проницаемости. Молекулы проходят через нее с различной скоростью: чем больше размер молекул, тем меньше скорость прохождения их через мембрану.
На внешней поверхности плазматической мембраны в животной клетке белковые и липидные молекулы связаны с углеводными цепями, образуя гликокаликс . Углеводные цепи выполняют роль рецепторов. Благодаря им осуществляется межклеточное узнавание. Клетка приобретает способность специфически реагировать на воздействия извне.
Под плазматической мембраной со стороны цитоплазмы имеются кортикальный слой и внутриклеточные фибриллярные структуры, обеспечивающие механическую устойчивость плазматической мембраны

 У растительных клеток кнаружи от мембраны расположена плотная структура - клеточная оболочка или клеточная стенка, состоящая из полисахаридов (целлюлозы)
У растительных клеток кнаружи от мембраны расположена плотная структура - клеточная оболочка или клеточная стенка, состоящая из полисахаридов (целлюлозы)
Схема строения клеточной стенки растений. О - срединная пластинка, / - первичная оболочка (два слоя по обе стороны от 0), 2 - слои вторичной оболочки, 3 - третичная оболочка, ПМ -
плазматическая мембрана, В - вакуоль, Я - ядро.
Компоненты клеточной стенки синтезируются клеткой, выделяются из цитоплазмы и собираются вне клетки, вблизи плазматической мембраны, образуя сложные комплексы. Клеточная стенка у растений выполняет защитную функцию , образует внешний каркас, обеспечивает тургорные свойства клеток. Наличие клеточной стенки регулирует поступление воды в клетку. Вследствие этого возникает внутреннее давление, тургор, препятствующее дальнейшему поступлению воды.